

Seed size variation impacts local adaptation and life-history strategies in a perennial grass
- PMID: 37132240
- PMCID: PMC10154936
- DOI: 10.1098/rspb.2022.2460
Seed size variation impacts local adaptation and life-history strategies in a perennial grass
Abstract
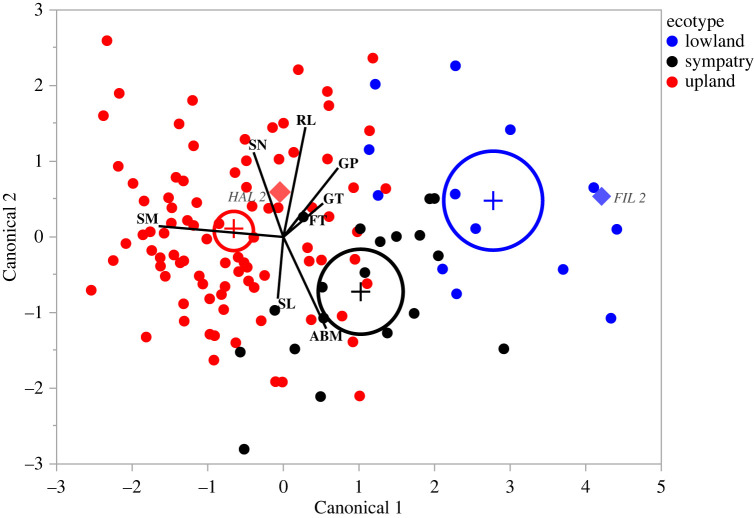
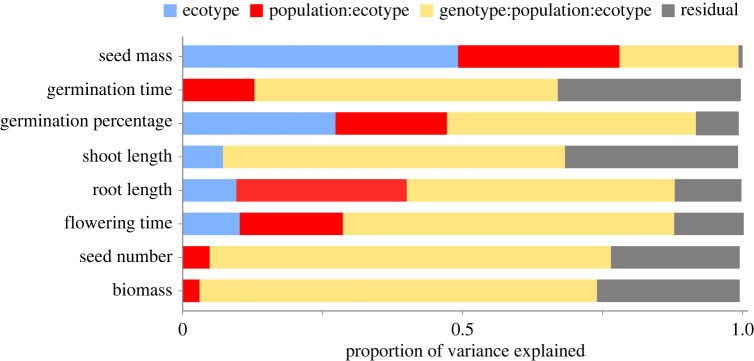
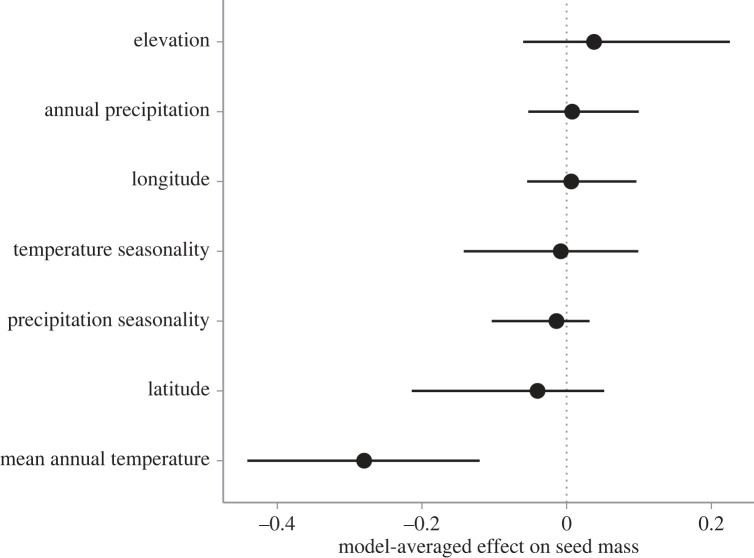
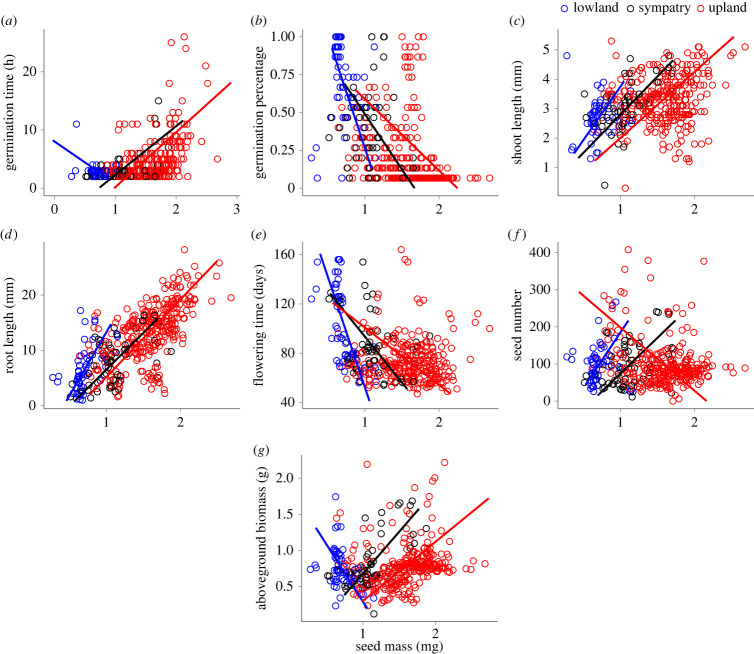
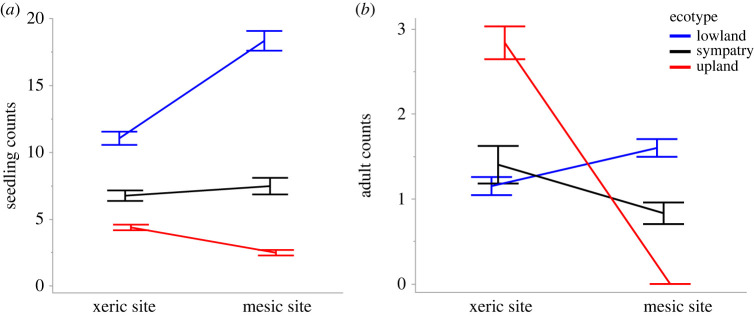
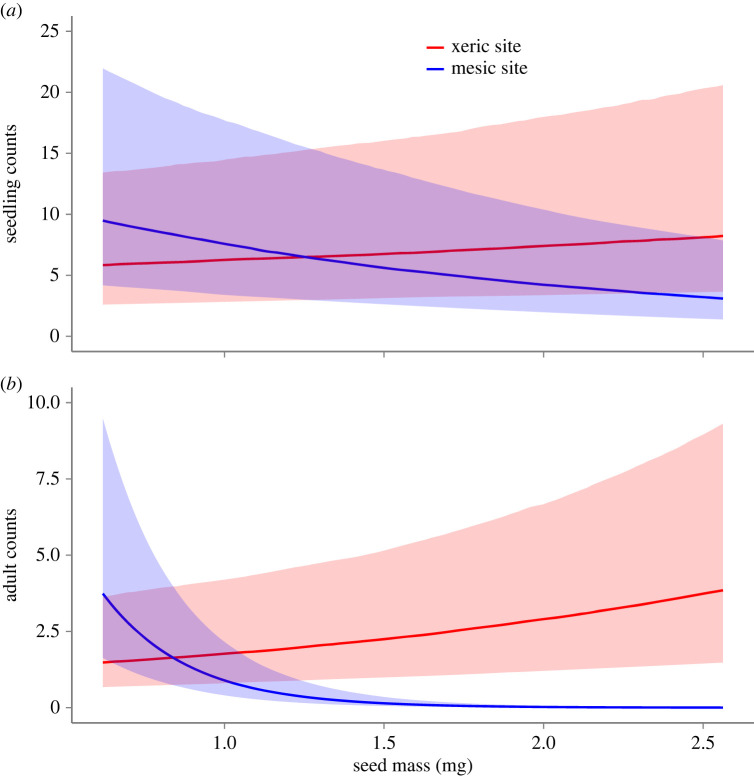
Seed mass is an ecologically important trait that often differs considerably among ecotypes. Yet, because few studies examine the impacts of seed mass on adult life-history traits, its role in local adaptation is unclear. In this study, using accessions of Panicum hallii that spanned the two major ecotypes, we examined whether covariation between seed mass, seedling and reproductive traits impacts ecotypic divergence and local adaptation. The perennial grass P. hallii has two distinct ecotypes-a large-seeded upland ecotype adapted to xeric environments and a small-seeded lowland ecotype adapted to mesic environments. In the greenhouse, seed mass varied greatly across P. hallii genotypes in a manner consistent with ecotypic divergence. Seed mass covaried significantly with several seedling and reproductive traits. At field sites representing the habitats of the two ecotypes, seed mass had different impacts on seedling and adult recruitment: selection favoured large seeds in upland habitat and small seeds in lowland habitat, which was consistent with local adaptation. By demonstrating the central role of seed mass in ecotypic differences in P. hallii and its importance to seedling and adult recruitment under field conditions, these studies show that early life-history traits can promote local adaptation and potentially explain ecotype formation.
Keywords: ecotypic differentiation; life-history evolution; local adaptation; trade-off.
Conflict of interest statement
We declare we have no competing interests.
Figures
Similar articles
-
The ecology and quantitative genetics of seed and seedling traits in upland and lowland ecotypes of a perennial grass.Evol Lett. 2022 Nov 22;6(6):460-473. doi: 10.1002/evl3.297. eCollection 2022 Dec. Evol Lett. 2022. PMID: 36579162 Free PMC article.
-
Seed traits and recruitment interact with habitats to generate patterns of local adaptation in a perennial grass.J Exp Bot. 2024 May 20;75(10):3111-3124. doi: 10.1093/jxb/erae063. J Exp Bot. 2024. PMID: 38381563
-
The genetics of divergence and reproductive isolation between ecotypes of Panicum hallii.New Phytol. 2015 Jan;205(1):402-14. doi: 10.1111/nph.13027. Epub 2014 Sep 23. New Phytol. 2015. PMID: 25252269 Free PMC article.
-
A Region on Chromosome 7 Related to Differentiation of Rice (Oryza sativa L.) Between Lowland and Upland Ecotypes.Front Plant Sci. 2020 Jul 27;11:1135. doi: 10.3389/fpls.2020.01135. eCollection 2020. Front Plant Sci. 2020. PMID: 32849696 Free PMC article. Review.
-
One hundred years into the study of ecotypes, new advances are being made through large-scale field experiments in perennial plant systems.Curr Opin Plant Biol. 2022 Apr;66:102152. doi: 10.1016/j.pbi.2021.102152. Epub 2022 Jan 19. Curr Opin Plant Biol. 2022. PMID: 35065527 Review.
Cited by
-
Variation in Seed Morphological Traits Affects the Dispersal Strategies of Chromolaena odorata Following Invasion.Plants (Basel). 2024 Jun 24;13(13):1747. doi: 10.3390/plants13131747. Plants (Basel). 2024. PMID: 38999587 Free PMC article.
References
-
- Grime JP, Hodgson JG, Hunt R. 2014. Comparative plant ecology: a functional approach to common British species. Berlin, Germany: Springer.
-
- Leishman MR, Westoby M. 1992. Classifying plants into groups on the basis of associations of individual traits: evidence from Australian semi-arid woodlands. J. Ecol. 80, 417-424. (10.2307/2260687) - DOI
-
- Venable DL. 1992. Size-number trade-offs and the variation of seed size with plant resource status. Am. Nat. 140, 287-304. (10.1086/285413) - DOI
-
- Moles AT, Westoby M. 2006. Seed size and plant strategy across the whole life cycle. Oikos 113, 91-105. (10.1111/j.0030-1299.2006.14194.x) - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
