

Chromatin Domains: The Unit of Chromosome Organization
- PMID: 27259200
- PMCID: PMC5371509
- DOI: 10.1016/j.molcel.2016.05.018
Chromatin Domains: The Unit of Chromosome Organization
Abstract
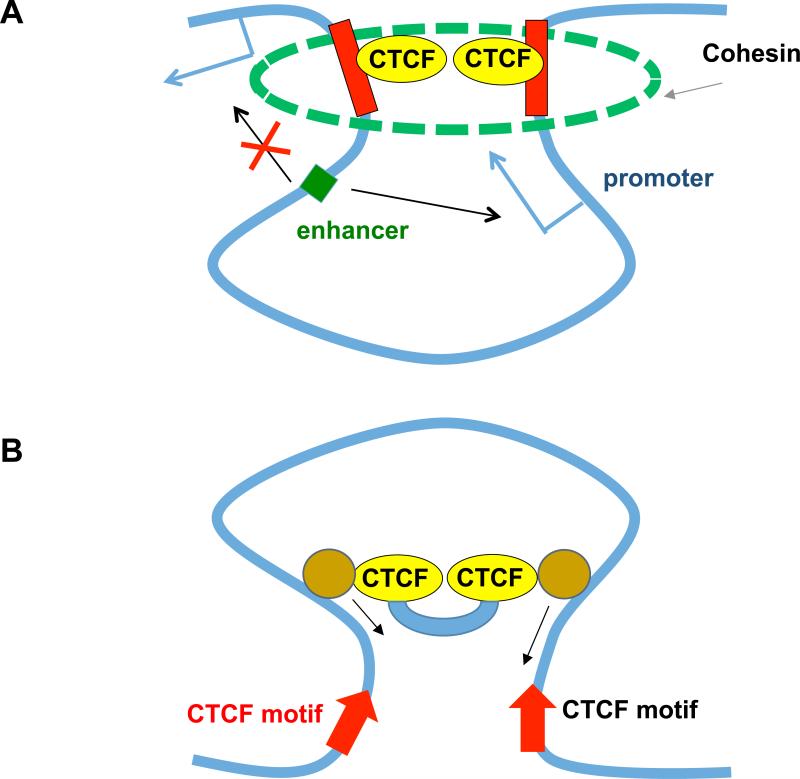
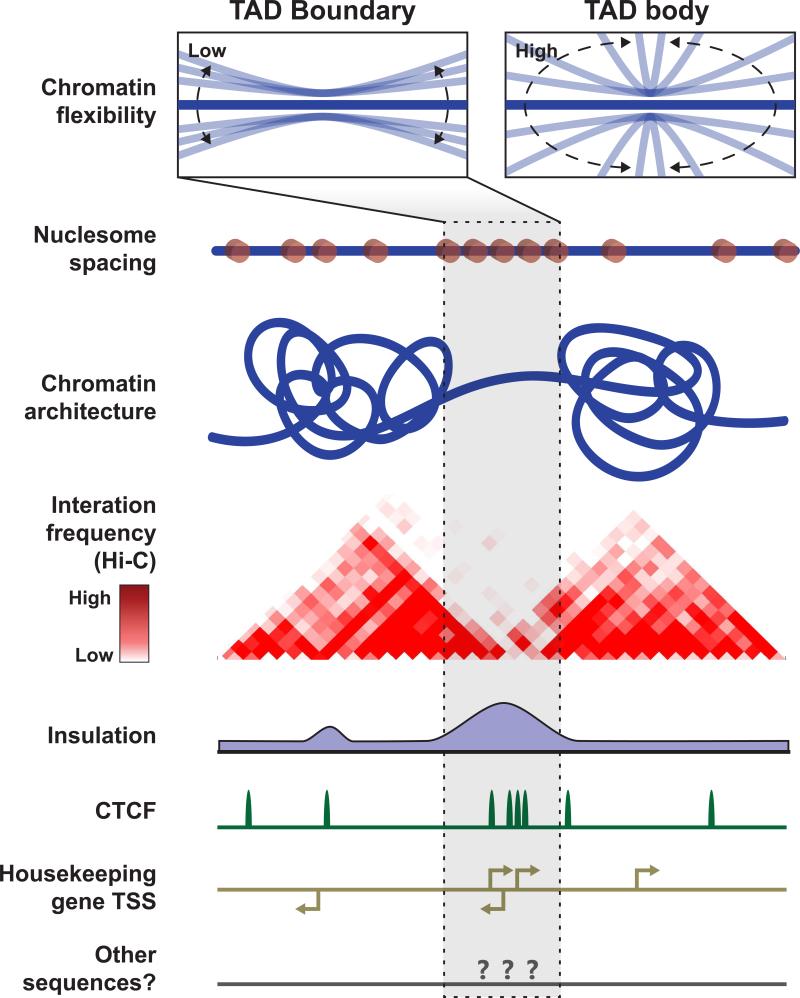
How eukaryotic chromosomes fold inside the nucleus is an age-old question that remains unanswered today. Early biochemical and microscopic studies revealed the existence of chromatin domains and loops as a pervasive feature of interphase chromosomes, but the biological implications of such organizational features were obscure. Genome-wide analysis of pair-wise chromatin interactions using chromatin conformation capture (3C)-based techniques has shed new light on the organization of chromosomes in interphase nuclei. Particularly, the finding of cell-type invariant, evolutionarily conserved topologically associating domains (TADs) in a broad spectrum of cell types has provided a new molecular framework for the study of animal development and human diseases. Here, we review recent progress in characterization of such chromatin domains and delineation of mechanisms of their formation in animal cells.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
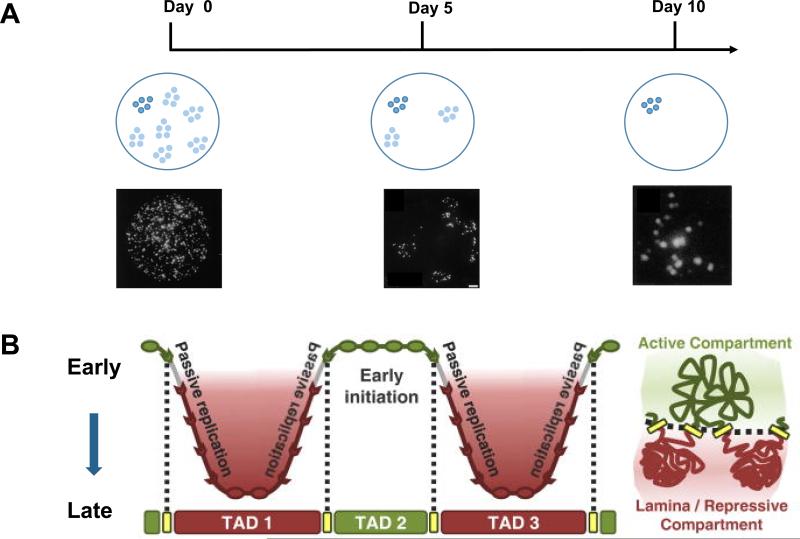
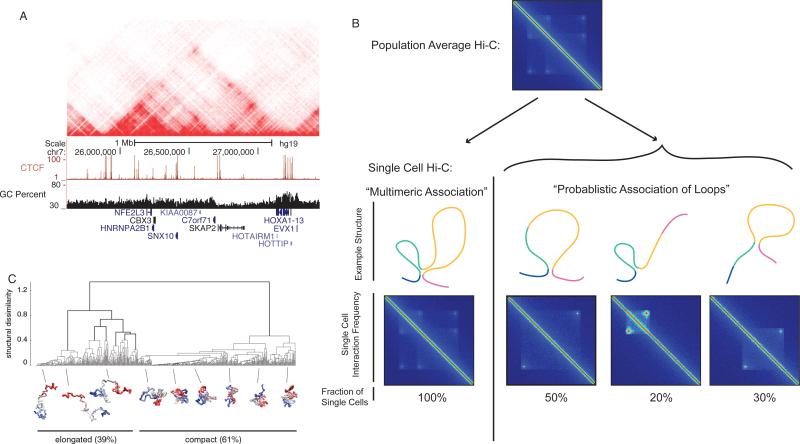
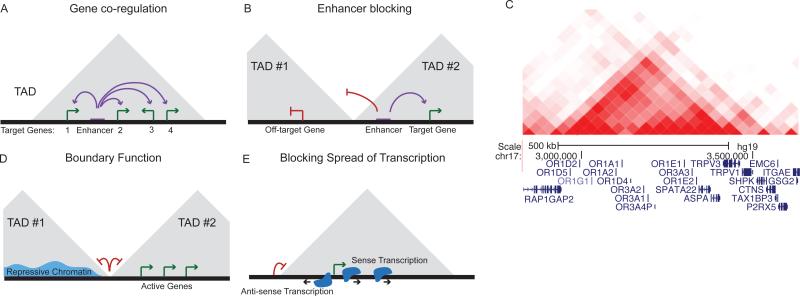
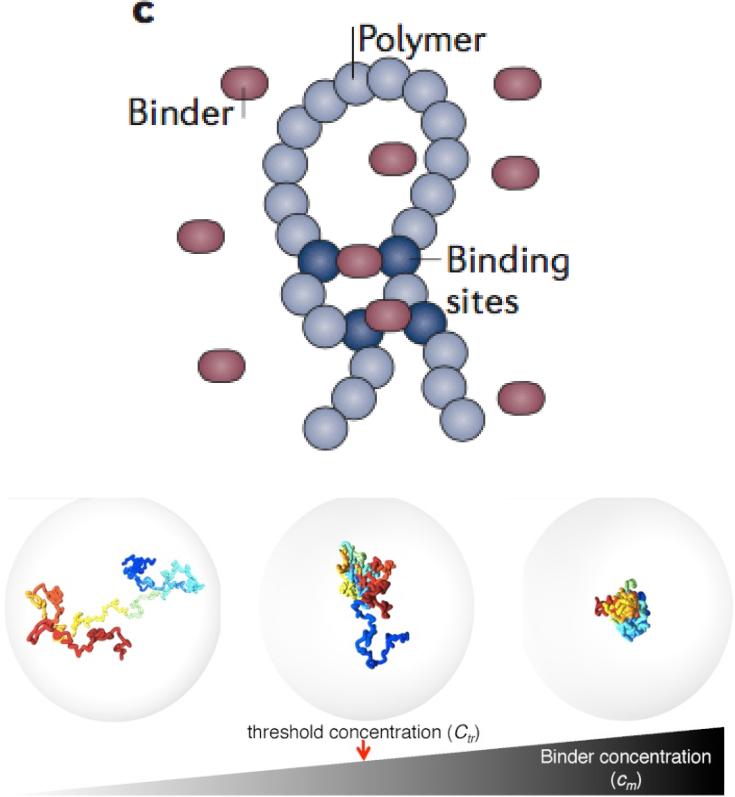
Similar articles
-
Spatial organization of chromatin domains and compartments in single chromosomes.Science. 2016 Aug 5;353(6299):598-602. doi: 10.1126/science.aaf8084. Epub 2016 Jul 21. Science. 2016. PMID: 27445307 Free PMC article.
-
Structural-Functional Domains of the Eukaryotic Genome.Biochemistry (Mosc). 2018 Apr;83(4):302-312. doi: 10.1134/S0006297918040028. Biochemistry (Mosc). 2018. PMID: 29626918 Review.
-
Principles of genome folding into topologically associating domains.Sci Adv. 2019 Apr 10;5(4):eaaw1668. doi: 10.1126/sciadv.aaw1668. eCollection 2019 Apr. Sci Adv. 2019. PMID: 30989119 Free PMC article. Review.
-
Chromatin folding--from biology to polymer models and back.J Cell Sci. 2011 Mar 15;124(Pt 6):839-45. doi: 10.1242/jcs.077628. J Cell Sci. 2011. PMID: 21378305
-
Chromatin Conformation Capture-Based Analysis of Nuclear Architecture.Methods Mol Biol. 2017;1456:15-32. doi: 10.1007/978-1-4899-7708-3_2. Methods Mol Biol. 2017. PMID: 27770354
Cited by
-
Integrating Multimorbidity into a Whole-Body Understanding of Disease Using Spatial Genomics.Results Probl Cell Differ. 2022;70:157-187. doi: 10.1007/978-3-031-06573-6_5. Results Probl Cell Differ. 2022. PMID: 36348107
-
Conformational state switching and pathways of chromosome dynamics in cell cycle.Appl Phys Rev. 2020 Sep;7(3):031403. doi: 10.1063/5.0007316. Appl Phys Rev. 2020. PMID: 32884608 Free PMC article.
-
3D genome organization contributes to genome instability at fragile sites.Nat Commun. 2020 Jul 17;11(1):3613. doi: 10.1038/s41467-020-17448-2. Nat Commun. 2020. PMID: 32680994 Free PMC article.
-
Processive DNA synthesis is associated with localized decompaction of constitutive heterochromatin at the sites of DNA replication and repair.Nucleus. 2019 Dec;10(1):231-253. doi: 10.1080/19491034.2019.1688932. Nucleus. 2019. PMID: 31744372 Free PMC article.
-
The MYC Enhancer-ome: Long-Range Transcriptional Regulation of MYC in Cancer.Trends Cancer. 2018 Dec;4(12):810-822. doi: 10.1016/j.trecan.2018.10.003. Epub 2018 Nov 2. Trends Cancer. 2018. PMID: 30470303 Free PMC article. Review.
References
-
- Andrey G, Montavon T, Mascrez B, Gonzalez F, Noordermeer D, Leleu M, Trono D, Spitz F, Duboule D. A switch between topological domains underlies HoxD genes collinearity in mouse limbs. Science. 2013;340:1234167. - PubMed
-
- Austenaa LM, Barozzi I, Simonatto M, Masella S, Della Chiara G, Ghisletti S, Curina A, de Wit E, Bouwman BA, de Pretis S, et al. Transcription of Mammalian cis-Regulatory Elements Is Restrained by Actively Enforced Early Termination. Mol Cell. 2015;60:460–474. - PubMed
-
- Bai XC, McMullan G, Scheres SH. How cryo-EM is revolutionizing structural biology. Trends Biochem Sci. 2015;40:49–57. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
