

Tests for the joint evolution of mating system and drought escape in Mimulus
- PMID: 21831854
- PMCID: PMC3278286
- DOI: 10.1093/aob/mcr160
Tests for the joint evolution of mating system and drought escape in Mimulus
Erratum in
- Ann Bot. 2012 Jun;109(7):1381
Abstract
Background and aims: Self-fertilizing taxa are often found at the range margins of their progenitors, where sub-optimal habitats may select for alternative physiological strategies. The extent to which self-fertilization is favoured directly vs. arising indirectly through correlations with other adaptive life history traits is unclear. Trait responses to selection depend on genetic variation and covariation, as well as phenotypic and genetic responses to altered environmental conditions. We tested predictions of the hypothesis that self-fertilization in Mimulus arises through direct selection on physiological and developmental traits that allow seasonal drought escape.
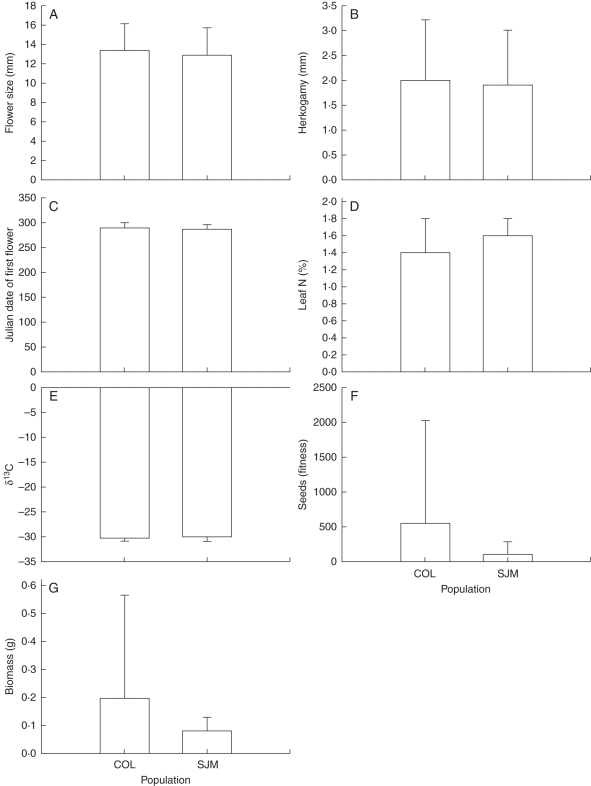
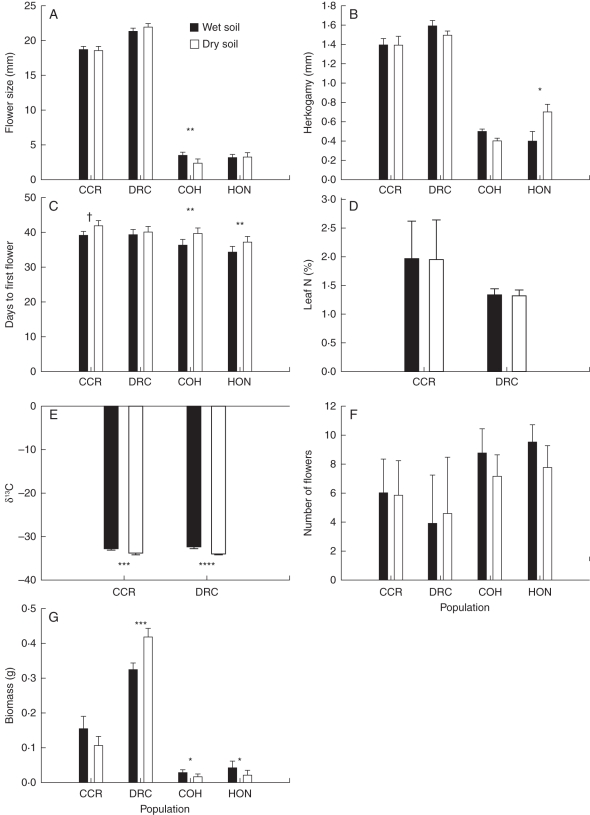
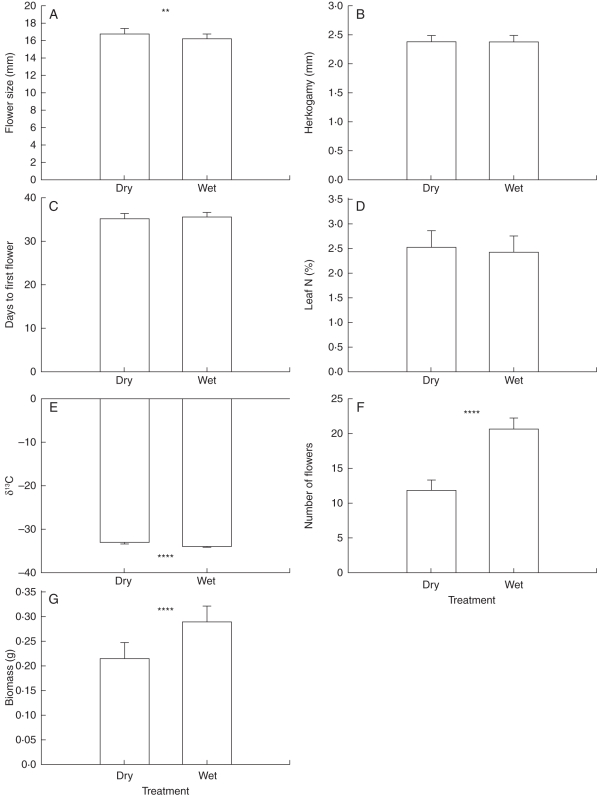
Methods: Phenotypic selection on mating system and drought escape traits was estimated in field populations of M. guttatus. In addition, trait phenotype and phenotypic selection were compared between experimental wet and dry soil in two greenhouse populations each of M. guttatus and M. nasutus. Finally, genetic variation and covariation for traits were compared between wet and dry soil treatments in a greenhouse population of M. guttatus.
Key results: Consistent with predictions, selection for early flowering was generally stronger than for mating system traits, and selection for early flowering was stronger in dry soil. Inconsistent with predictions, selection for water-use efficiency was largely absent; selection for large flowers was stronger than for drought escape in the field; and most drought escape and mating system traits were not genetically correlated. A positive genetic correlation between flowering time and flower size, which opposed the adaptive contour, emerged only in wet soil, suggesting that variation in water availability may maintain variation in these traits. Plastic responses to soil moisture treatments supported the idea that taxonomic divergence could have been facilitated by plasticity in flowering time and selfing.
Conclusions: The hypothesis that plant mating systems may evolve indirectly via selection on correlated life history characteristics is plausible and warrants increased attention.
Figures
Similar articles
-
Replicate altitudinal clines reveal that evolutionary flexibility underlies adaptation to drought stress in annual Mimulus guttatus.New Phytol. 2015 Apr;206(1):152-165. doi: 10.1111/nph.13153. Epub 2014 Nov 18. New Phytol. 2015. PMID: 25407964
-
Minor quantitative trait loci underlie floral traits associated with mating system divergence in Mimulus.Evolution. 2002 Nov;56(11):2138-55. doi: 10.1111/j.0014-3820.2002.tb00139.x. Evolution. 2002. PMID: 12487345
-
Could seasonally deteriorating environments favour the evolution of autogamous selfing and a drought escape physiology through indirect selection? A test of the time limitation hypothesis using artificial selection in Clarkia.Ann Bot. 2018 Mar 14;121(4):753-766. doi: 10.1093/aob/mcx197. Ann Bot. 2018. PMID: 29351591 Free PMC article.
-
The genetic control of flower-pollinator specificity.Curr Opin Plant Biol. 2013 Aug;16(4):422-8. doi: 10.1016/j.pbi.2013.05.004. Epub 2013 Jun 10. Curr Opin Plant Biol. 2013. PMID: 23763819 Free PMC article. Review.
-
The evolution of drought escape and avoidance in natural herbaceous populations.Plant Sci. 2015 May;234:155-62. doi: 10.1016/j.plantsci.2015.02.012. Epub 2015 Feb 25. Plant Sci. 2015. PMID: 25804818 Review.
Cited by
-
Experimental evolution suggests rapid assembly of the 'selfing syndrome' from standing variation in Mimulus guttatus.Front Plant Sci. 2024 Aug 27;15:1378568. doi: 10.3389/fpls.2024.1378568. eCollection 2024. Front Plant Sci. 2024. PMID: 39263417 Free PMC article.
-
GIGANTEA Unveiled: Exploring Its Diverse Roles and Mechanisms.Genes (Basel). 2024 Jan 13;15(1):94. doi: 10.3390/genes15010094. Genes (Basel). 2024. PMID: 38254983 Free PMC article. Review.
-
The influence of genetic architecture on responses to selection under drought in rice.Evol Appl. 2022 Jun 6;15(10):1670-1690. doi: 10.1111/eva.13419. eCollection 2022 Oct. Evol Appl. 2022. PMID: 36330294 Free PMC article.
-
Genetic and spatial variation in vegetative and floral traits across a hybrid zone.Am J Bot. 2022 Nov;109(11):1780-1793. doi: 10.1002/ajb2.16067. Epub 2022 Oct 4. Am J Bot. 2022. PMID: 36193908 Free PMC article.
-
Multivariate selection mediated by aridity predicts divergence of drought-resistant traits along natural aridity gradients of an invasive weed.New Phytol. 2022 May;234(3):1088-1100. doi: 10.1111/nph.18018. Epub 2022 Feb 28. New Phytol. 2022. PMID: 35118675 Free PMC article.
References
-
- Angert AL. Growth and leaf physiology of monkeyflowers with different altitude ranges. Oecologia. 2006;148:183–194. - PubMed
-
- Armbruster WS, Mulder CPH, Baldwin BG, Kalisz S, Wessa B, Nute H. Comparative analysis of late floral development and mating-system evolution in tribe Collinsieae (Scrophulariacae) American Journal of Botany. 2002;89:37–49. - PubMed
-
- Arntz AM, Delph LF. Pattern and process: evidence for the evolution of photosynthetic traits in natural populations. Oecologia. 2001;127:455–467. - PubMed
-
- Arntz AM, DeLucia EH, Jordan N. Fitness effects of a photosynthetic mutation across contrasting environments. Journal of Evolutionary Biology. 2000;13:792–803.
-
- Arroyo MTK. Chiasma frequency evidence on the evolution of autogamy in Limnanthes floccosa (Limnanthaceae) Evolution. 1973;27:679–688. - PubMed
